|
||||
 |
|||||
Major research topics in Nikilab |
|||||
<Chromosome Segregation in Bacteria> |
|||||
Chromosome must be precisely segregated into daughter cells for bacterial cells to proliferate. Although prokaryotic cells does not have “nuclei” that exist in Eukaryote, the chromosomal DNA, which is about 1000 times longer than cell length, is tightly condensed as “nucleoid” within the cell. |
|||||
|
1. Centromere Like Region in Bacteria |
||||
|
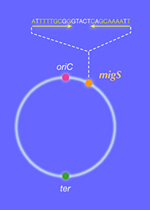
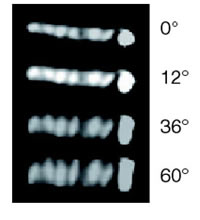
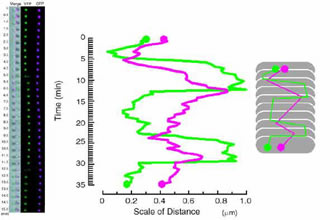
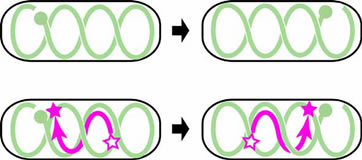
For long years, Chromosome segregation in Bacteria had been thought to be a passive event that chromosomal DNA migrates toward daughter cells according to extension of the cell membrane to which the origins of chromosomes are anchored (Replicon Model of Jacob, F. et al., 1963). Recent study, however, revealed that specific region on chromosome of E. coli can dynamically migrate toward cell poles independently of the extension of the cell membrane. On the contrary to the eukaryotic cells in which each step of replication and segregation of chromosomal DNA are apparently distinguishable, in bacteria, specific region of chromosomal DNA migrates toward cell poles and partitioned during replication. In this migration event, repliucation origin, oriC and terminal, ter shows different dynamics. Replication origin regions migrate to cell poles promptly after its replication and stay there till cell division. On the other hand, ter region stayed still at mid cell until just before cell division. Is there a centoromere like region at replication origin in former case? We revealed that the origin region migrates to cell poles via “migS” region on chromosome (Fig1, Yamaichi, Y. and Niki, H., 2004). Further more the chromosome DNA showed dynamic structural change (Fig. 2???). Now we are exploring and analyzing the proteins that form complex with the migS region. |
||||
2. A moter protein that moves DNA |
|||||
|
To migrate the replicated DNA toward cell poles as described above, there must exist driving force produced by motor protein or other factor. Recently, ATPase proteins, which are essential for stable partitioning of low copy number plasmids, are focused as the motor protein for DNA segregation.
|
||||
< Novel Genetic System for Chromosome Study > Yeasts are powerful genetic tool to identify new elements or new phenotypes in a wide range of biology. However, due to its small cell size, there is a limitation in detailed cell biological analysis. Schizosaccharomyces japonicus is an alternative fission yeast. Although this haploid organism is poorly developed as a genetic tool, its nearly twice large nuclear size and fibrously condensed mitotic chromosomes compare to the other yeasts can be more suitable model system to understand mechanism of chromosome organization in vivo, such as mitotic chromosome segregation or interphase chromosome compartment. Our final aim is to discover new mechanisms that regulate chromosome cycle through the isolation of mutants with novel cytological phenotypes. We are currently collecting mutants showing disorganized chromosome behaviors. In parallel, we are generating chromosomal markers to ease our analysis. Thanks to Broad Institute, S. japonicus genomic sequences are fully available, and we have already tagged several important factors to visualize those components.
|
|||||
国立遺伝学研究所 〒411-8540 静岡県三島市谷田1111-1 > 連絡先詳細および地図はこちら |
Microbial Genetics Laboratory (Niki Laboratory) 1111-1 Yata Mishima Shizuoka Japan P.O. 411-8540 > Contact details and Maps |